the different varietals in Africa
the gene map

Coffea arabica allotetraploidy resulted from recent natural hybridization between the ancestors of present‐day C. canephora (Ca genome donor) and C. eugenioides (Ea genome donor) (Lashermes et al ., 1999) and is probably the result of a single event (Lashermes et al ., 2014). Coffea arabica displays disomic inheritance with bivalent pairing of homologous chromosomes (Krug and Mendes, 1940), which is in accordance with our present observations in mapping experiments of usual disomic patterns of marker inheritance.
Yu et al . (2011) provided evidence of recent C. arabica speciation no more than 0.665 million years ago, but also low divergence between the two constitutive subgenomes of C. arabica (Ca and Ea) and those of its progenitor species, demonstrating that the nuclear genomes have remained essentially unaltered since the formation of the hybrid.
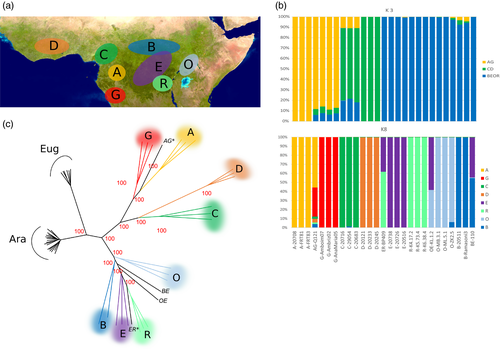
Using the 17 C. arabica accessions, we took advantage of the fact that they exhibited close genetic relationships in order to apply a haploid‐genotype‐based assignation procedure to discover its closest current relatives in our sampling. Using our Coffee8.5K genotyping, we could not preferentially associate the Ea subgenome to any of the sampled individuals (Figure 4d).
On the contrary, we were able to preferentially assign the Ca subgenome to C. canephora of the O (Ugandan) Diversity group (Figure 3) and the shortest distance was observed with a North Ugandan C. canephora individual from the Zoka Forest. Geographically, this forest contains the closest C. canephora population to the current C. arabica distribution (Figure 4a–c).
At the present time, C. arabica is mainly found in the southwestern highlands of Ethiopia (Figure 4a), with some occurrence on the Boma plateau in southeastern South Sudan (Thomas, 1942), and on Mount Imantong in Sudan and Mount Marsabit in northern Kenya (Berthaud and Charrier, 1988). C. arabica is the main Coffea species that occurs in those regions and is geographically isolated from all diploid coffee species in the genus, which includes its two progenitor species C. canephora and C. eugenioides . Moreover, C. arabica also differs from C. canephora in terms of current environmental requirement and predicted niche distribution (Gomez et al ., 2016). Thus, precise localization in Africa of the cradle of C. arabica , based on the present distribution of its two progenitor species appears difficult.
Two non‐exclusive scenarios could be suggested for the origin and geographical isolation of C. arabica . (i) Upon hybridization, C. arabica could have followed one of the typical patterns of the polyploid distribution with peripheral expansion outside the range of the distribution of its diploid parental species. Indeed, it has been suggested that plants with double genomes—auto‐ or allo‐polyploids—have the potential to develop phenotypic novelties, increase their adaptability and obtain higher fitness features that would render them more tolerant towards changing conditions than their diploid counterparts (Amborella Genome et al ., 2013). This was particularly well illustrated with the survival and proliferation of polyploid plant lineages during the Cretaceous–Tertiary mass extinction event (Fawcett et al ., 2009). Following the hypothesis that hybridization may favour establishment in novel habitats (Pillon et al ., 2009; Rieseberg et al ., 2007), C. arabica could have migrated northwards—away from the overlapping distribution range of C. canephora and C. eugenioides— towards its current distribution range with specific environmental requirement (Gomez et al ., 2016). Its original population is genetically related to the current Zoka population, which thus represents the southernmost remnant of the original distribution. (ii) Meanwhile, considering that the constitution and the extent of tropical forests have varied considerably during the late Quaternary period (Mumbi et al ., 2008), past C. canephora and C. eugenioides distributions may have been more widespread in higher latitudes and could have overlapped with the current C. arabica distribution in habitats that were suitable for all three species. In fact, C. canephora was probably able to find suitable habitats in Ethiopia in the recent past (mid‐Holocene, ~6000 years before present, R. Tournebize, data not shown). The climatic changes could have reduced the diploid distributions to their current locations. As a consequence, the birthplace of C. arabica could possibly be not only in Ethiopia but also in the entire region (South Sudan, Uganda, North Kenya) followed either by migration to present‐day Ethiopia or by survival in that region alone.
Due either to past habitat shift in Eastern Africa or to the colonization of new peripheral geographical areas, the presence of C. arabica in different environments and a reduction in diploid competitors would have increased the divergence of initially con‐specific populations and eventually would have given rise to the C. arabica speciation. C. arabica's self‐fertility would have further contributed to its genetic isolation.

wiley online
留言
張貼留言